在熱帶淺海中,生活著一種體長僅 5 毫米的小蝦 – 少刺透鏡糠蝦。雖然牠們的身影微小,但牠們那直徑僅 0.4 毫米的眼球裡,卻藏著自然界最精妙的光學工程奇蹟。

複眼的極致演化:從複眼到高解析「單眼」
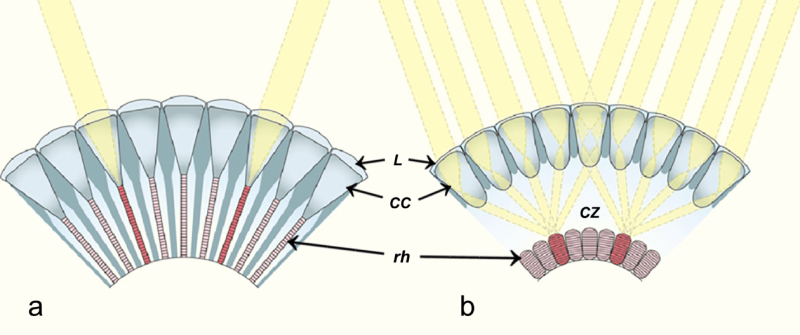
大多數甲殼類動物的眼睛屬於複眼,由成百上千個稱為小眼 (ommatidia) 的單元組成。在糠蝦這類動物中,牠們普遍採用的是疊置型複眼 (superposition eye)。這種構造非常巧妙:在小眼外部的光學單元(透鏡與晶錐)與底部的感光單元(感桿束)之間,存在一個被稱為「透明區」 (clear zone) 的空間。這種配置可以增加「集光效率」,來自空間中同一個點的光線,穿過複眼表面多個不同的小眼,經過光學單元的折射,再經過透明區,最終會匯聚在視網膜上的同一個點。這種設計極大化了集光能力,讓動物能在昏暗的水下環境中仍保有視覺。
然而,複眼有一個致命的弱點:解析度低。在光學中,繞射 (diffraction) 限制了影像的清晰度,而繞射現象與鏡片的直徑成反比。複眼由成百上千個微小的小眼組成,每個小眼直徑極小(如糠蝦的普通小眼僅 16 微米)。這導致單個小眼的繞射模糊非常嚴重。如果複眼想要提升一倍的解析度,根據計算,其小眼面 (facet) 直徑必須增加一倍,這會導致眼睛整體的表面積以幾何倍數暴增。對於體長僅 5 毫米的糠蝦來說,牠的大腦與身體根本裝不下那樣巨大的複眼。
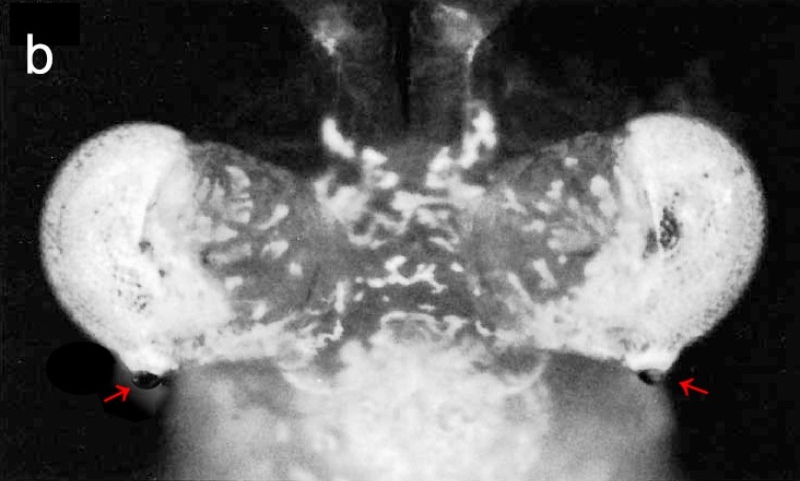
面對物理限制,少刺透鏡糠蝦演化出一種局部特化的新奇策略,形成一個高解析區 (acute zone)。少刺透鏡糠蝦最引人注目的特徵,是鑲嵌在每隻複眼後側的一個巨大小眼。從上方觀察,這個巨大的小眼向後凸出,對著後方的景物。
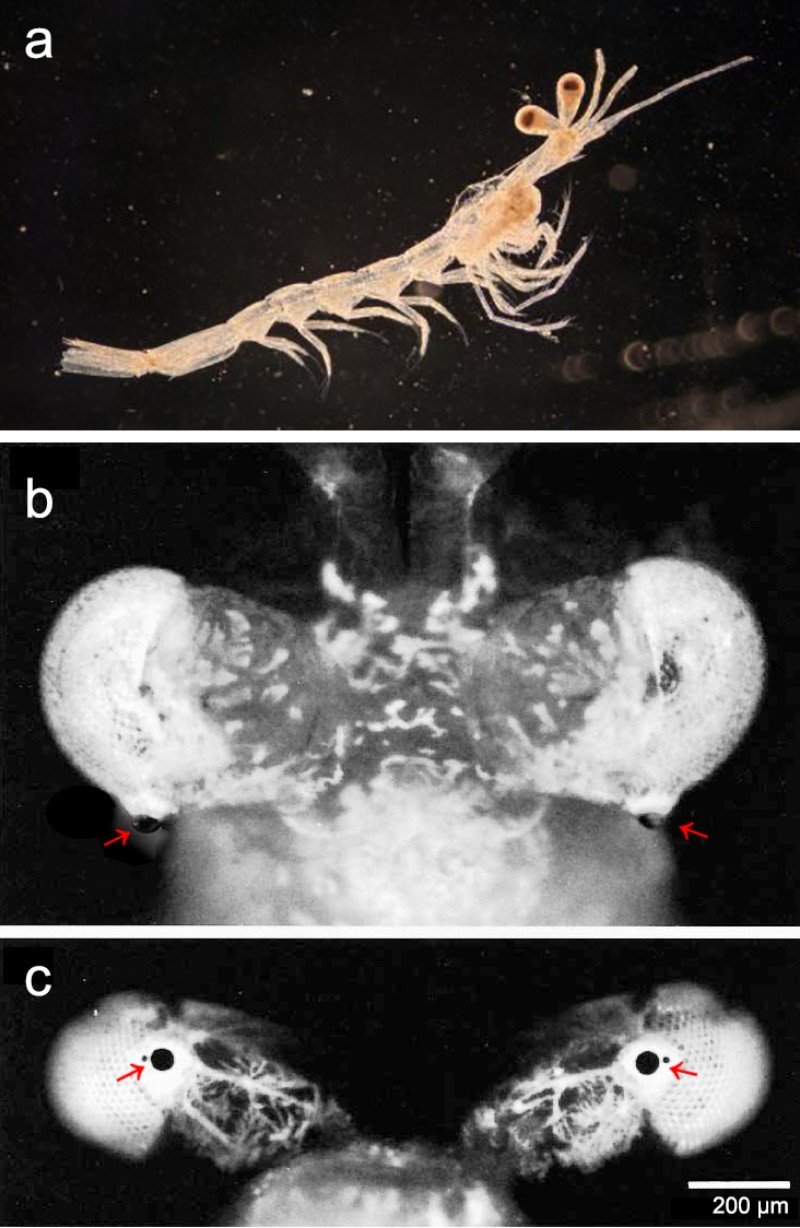
科學家透過組織切片發現,這個巨大的小眼有著巨大的透鏡,直徑是普通小眼的 3 倍,透鏡下方有一個巨型晶錐,而晶錐下方竟排布著 120 個極其細長的感桿束 (rhabdoms)。在典型的複眼中,每個小眼僅對應一個感桿束;因此可以推估,這塊區域原本是由 120 個獨立小眼所組成。為了追求更好的解析度,演化讓其中絕大部分的小眼放棄了自己的「鏡頭」(透鏡與晶錐),僅保留最核心的感光單元(感桿束),並將其壓縮、拉長,共同緊密排列成一片特化的視網膜陣列。最終,這 120 個感光單元共享同一個由特化小眼發育而成的巨型鏡頭。這種「一鏡對多點」的特殊構造,本質上等於將局部的複眼區改造為一個高解析度的「單眼」。不同於正常複眼區是無數的小眼各自為政,這顆特化的單眼的運作模式更像人類的相機眼:由單一巨大鏡頭捕捉光線,並將高解析影像投射到下方由 120 個感光像素構成的「微型視網膜」上。
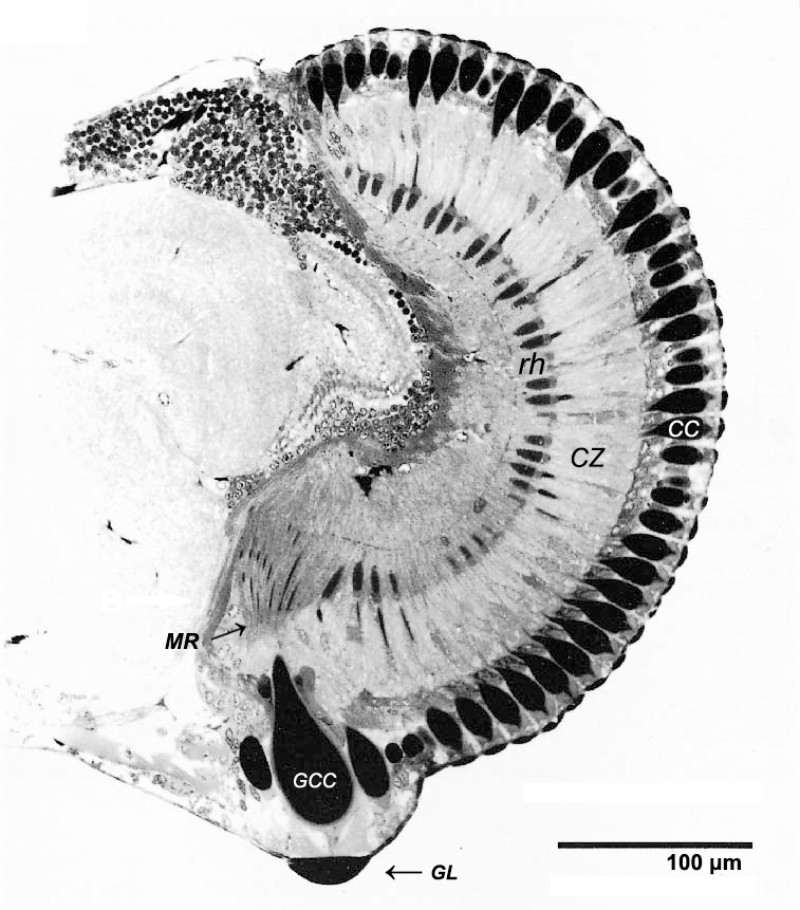
從構造到性能:微型世界的「光學升級」
這種結構上的劇變,直接轉化為性能上的飛躍。根據 Nilsson 等人的計算,這塊被稱為「高解析區」的特殊構造,其解析度比周圍正常的複眼區高出 6.4 倍。雖然這 120 個像素在數量上遠不及現代數位相機,但在僅 0.4 毫米的眼球尺度下,這已是接近光學極限(繞射極限)的極致表現。
然而,這種「高清底片」也帶來了副作用;由於光線被分散到 120 個感光點,單個像素接收到的光量大幅減少,導致感光靈敏度下降。這意味著這套系統無法像普通的疊置複眼那樣在深海微光中運作,而是專為日間、強光下的精準辨識而設計。
視覺模式的切換:從後方收納到前方鎖定
少刺透鏡糠蝦在自然棲息的狀態下,牠的高解析鏡頭是指向身體後側水平線上方約 12° 的方向。涵蓋了 15 – 20° 的視野,雙眼重疊區大,能形成立體視覺;因此糠蝦可透過左右眼觀察角度的微小差異,辨識出標的物的距離。對於需要尋找獵物或避開掠食者的動物來說,將「最強鏡頭」對準後方似乎並不符合邏輯。
然而,從效益最大化的角度來看,這其實是很合理的空間配置。由於高解析區的感光靈敏度極低(僅普通小眼區的 1/10)。如果這對「單眼」位於正前方,將會導致糠蝦在低光環境下出現嚴重的「前方盲區」。因此,平時將這對高清鏡頭向後「收納」,能確保正常的複眼擁有完整且無遮蔽的前方視野,用於偵測大範圍的威脅。那麼糠蝦如何運作這對高解析「單眼」呢?研究發現,少刺透鏡糠蝦具備令人驚嘆的眼球靈動性。首先,在水平面上,牠們的眼柄可以向左右擺動各60°,眼柄本身更可以旋轉達 130°。這意味著原本指向後方的「高解析鏡頭」,除了可以向左右調整方向,還可以轉向上方或前方。再者,牠們左右眼的轉動是完全同步的。無論眼柄如何轉動,這一對高解析鏡頭始終對著同一個方向。平時,高解析區處於「休息」位置指向後方,這可能是為了讓前方的視覺場保留給靈敏度較高、視野較廣的普通小眼區,以便偵測大範圍的威脅與導航。一旦發現潛在標的,牠們會迅速轉動眼柄,切換到「鎖定模式」,利用高解析鏡頭監測目標。
演化上的奇觀
少刺透鏡糠蝦的視覺系統,是自然界應對光學難題的絕佳範例。即便在數毫米的微小空間內,演化仍能透過精巧的結構重組與行為配套,展現生命的高度複雜性。這讓人聯想到 3 億 9 千萬年前泥盆紀海洋中的鏡眼三葉蟲。2021 年的研究指出,這類古生物演化出了獨特的「超複眼」系統—在其巨大的裂色眼透鏡下方,竟然隱藏著由微小視覺單元組成的次級複眼結構。從遠古三葉蟲的「超複眼」到今日糠蝦的局部「單眼」,自然界一再地在複眼架構上,透過局部特化與結構疊加來突破生理極限,演繹出超越想像的視覺創新。

延伸閱讀
- Nilsson DE & Modlin RF 1994. A MYSID SHRIMP CARRYING A PAIR OF BINOCULARS. The Journal of Experimental Biology, 189 1, 213-36.
- 三億九千萬年前的奇蹟之眼:鏡眼蟲的超複眼構造
- 三葉蟲捲曲身體的防禦策略
- 消失的古生物!由三葉蟲眼睛構造的研究,支持其親緣歸屬是有顎類的姊妹群
- 能用腳壓碎獵物外殼的三葉蟲

本著作由本館研究人員所提供,博學多文團隊編輯製作,以創用CC 姓名標示–非商業性– 禁止改作 4.0 國際 (CC BY-NC-ND 4.0) 授權條款釋出。若需要使用本篇的文字、圖像等,請洽本館出版室。