三葉蟲及一些早期節肢動物具有和現生鱟同源的附肢結構,科學家比對了兩種三葉蟲與鱟的附肢構造,推測其中的霸王萊德利基蟲具有和鱟類似的攝食機制,能用附肢上的顎基將帶殼獵物壓碎後再送進嘴裡,可說是最早發現的食殼性動物。食殼性的出現可能也帶動了掠食者與有殼獵物的軍備競賽。

幫助攝食的附肢
節肢動物會用附肢來協助攝食,如有顎類 (Mandibulata) 的螃蟹會用螯將食物碎屑送入口中,螳螂會用前足攫取獵物慢慢享用;而螯肢類 (Chelicerata) 的蠍子也會用鉗肢固定獵物,蜘蛛會用特化的螯肢將毒液注入獵物體內,其中著名的活化石—鱟則會用口後方附肢基節上的顎基將有殼獵物壓碎後再「咀嚼」送入口中。
三葉蟲的食性
三葉蟲是古生代最成功的節肢動物類群,牠們廣布於古生代的海洋中,從熱帶淺海、珊瑚礁,到極地深海,涵蓋了不同的緯度以及深度,各有不同的生態區位,且形態多樣,顯示牠們也有多樣的生活模式及營養(攝食)類型,包括了掠食、食腐、濾食、食浮游生物、寄生等,甚至有些三葉蟲是化學自營共生的宿主。但一般認為最早的三葉蟲是掠食者。雖然近年有研究認為三葉蟲與有顎類是姊妹群,但長久以來,鱟(螯肢類)一直被認為是與三葉蟲親緣最近的現生生物,因此兩者間也常被拿來做比對。科學家選擇了兩種保存極好的寒武紀三葉蟲,一是澳洲鴯鶓灣頁岩的霸王萊德利基蟲,另一是伯吉斯頁岩的鋸齒擬油櫛蟲。藉由比較這些三葉蟲與現生鱟的附肢,推測其中的霸王萊德利基蟲會用附肢將帶殼獵物壓碎後再送進嘴裡。這種能掠食具有堅硬外殼獵物的食性稱食殼性。
節肢動物的食殼性
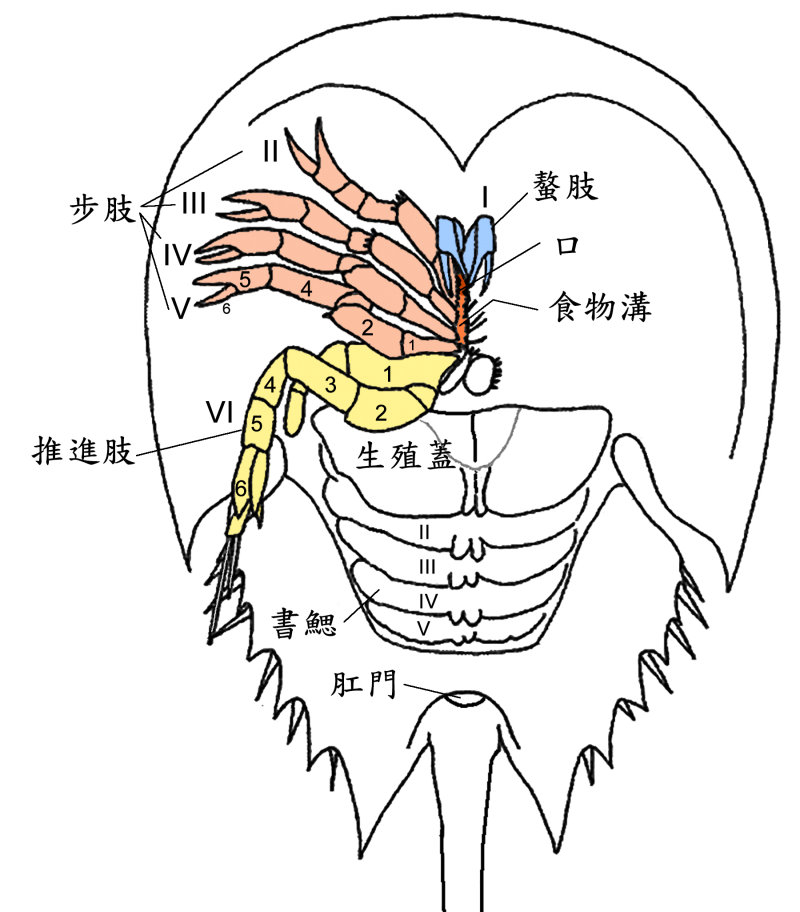
食殼性是有效的攝食模式。食殼的生物會先壓碎獵物的角質或硬殼,例如貝殼,再把牠吃進去。現生的節肢動物-美洲鱟即具有這種特化功能,牠會用附肢壓碎有殼獵物。從腹面看,鱟有六對附肢,其第一對是螯肢(I), II-V 是步肢,VI 是推進肢。附肢 II-VI 的近端第一節稱為基節 (coxa),基節的內側有特化的帶刺顎基 (gnathobase),兩兩相對的顎基共同形成鱟處理食物的食物溝。鱟用附肢 II-IV 上的顎基來「咀嚼」獵物,因為這些顎基上的棘較長而精緻,可以撕碎較軟的食物。而附肢 V 和 VI 的顎基刺粗壯,則可以用來壓碎獵物的殼體。這種顎基咀嚼系統由一系列基本上同質的運動/攝食附肢組成,在古生代的節肢動物中很常見。
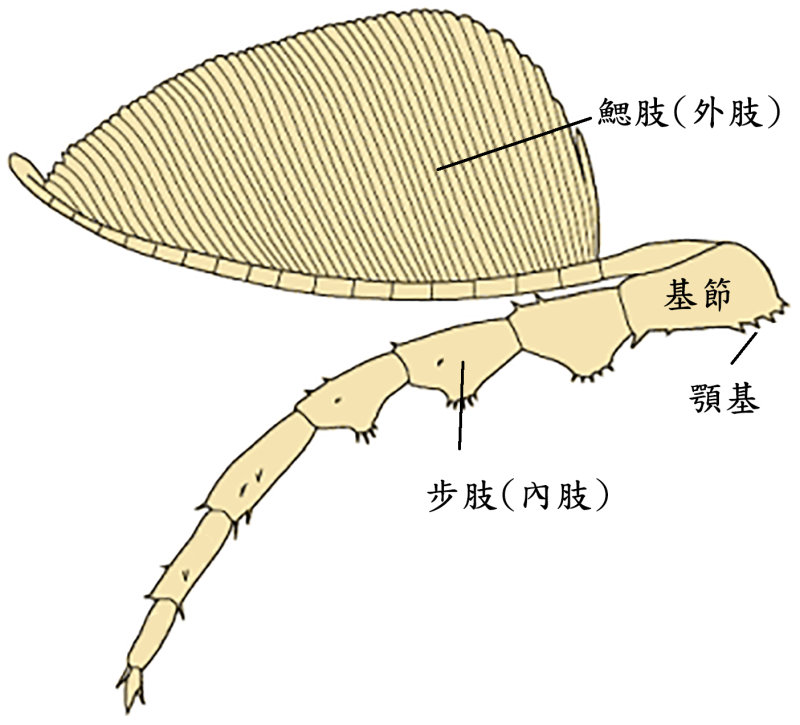
食殼性三葉蟲
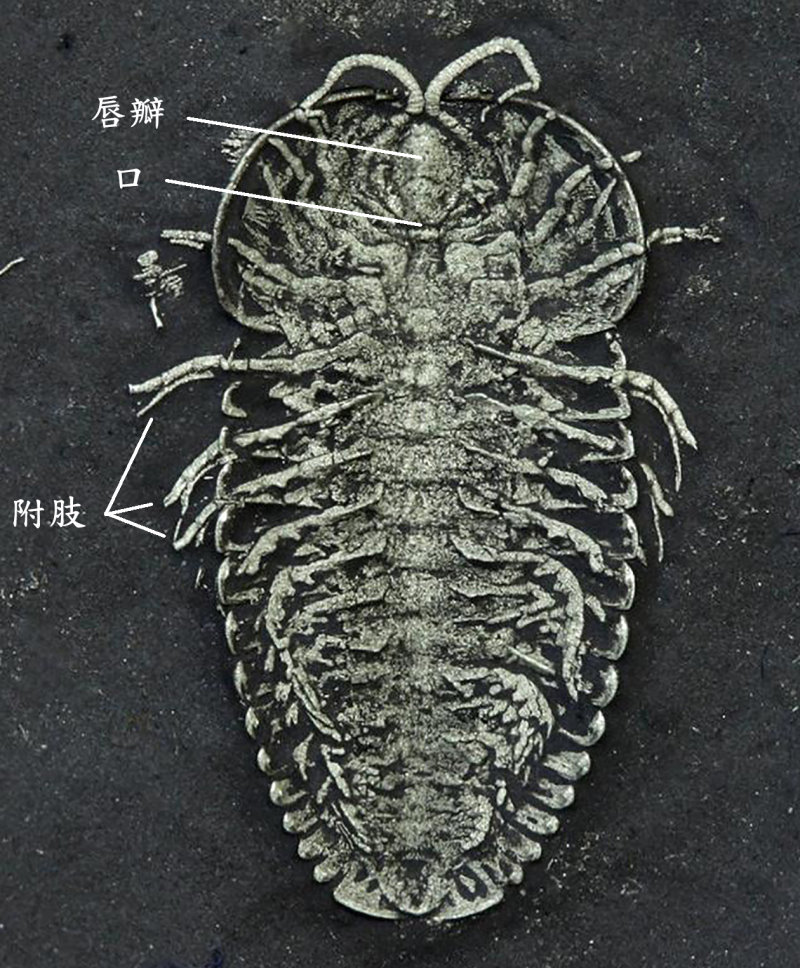
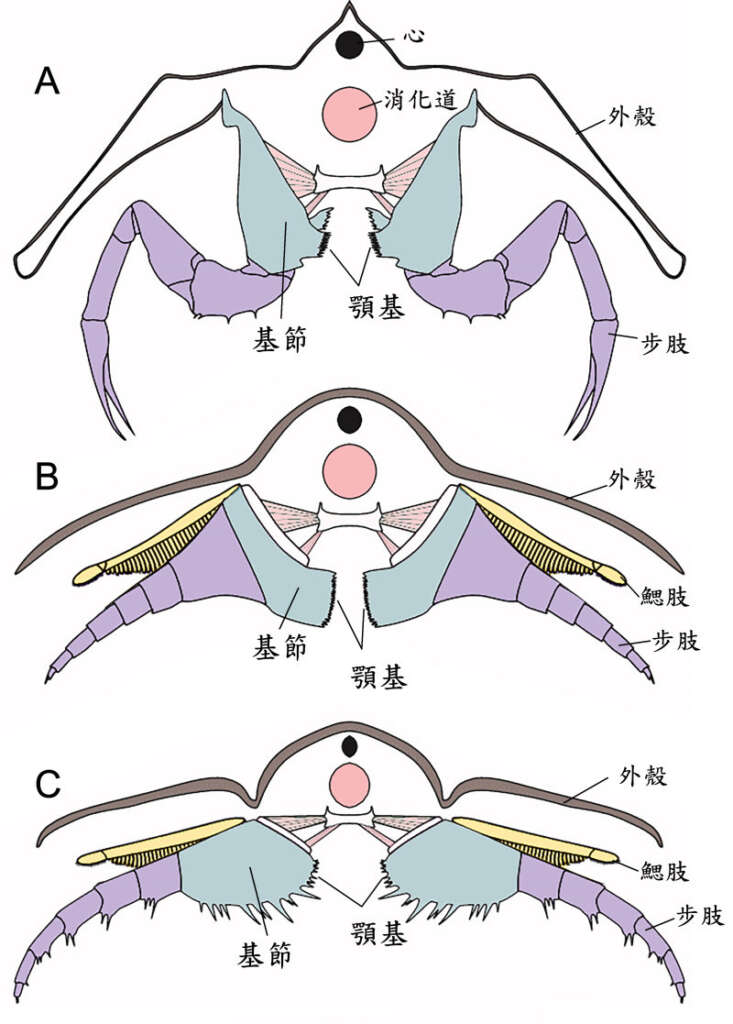
三葉蟲頭部的腹側有一個突起的唇瓣,唇瓣後端即為口,口部之後是成對的附肢,在左右兩排附肢的基節之間,即是可與鱟構造對應的食物溝。三葉蟲的附肢是雙肢型,在上方的分支稱外肢(鰓肢),具呼吸功能;下方的稱內肢(步肢),具運動功能。三葉蟲的步肢具有與鱟同源的基節,且基節上亦有帶刺的顎基構造。科學家根據保存精緻的化石復原了鋸齒擬油櫛蟲和霸王萊德利基蟲的附肢模型。並以鱟的同源肌群推測這些已滅絕的生物附肢基節上的應力分布。下圖 A 是鱟的附肢 V 的前視縱切面,可見其基節上的顎基相對向內,黑色區域為其分布的短刺,此即鱟處理硬殼的地方,壓碎獵物的殼後,即往前送,用附肢 IV-II 的顎基來進一步處理,並將食物送入口中。下圖 B 的霸王萊德利基蟲的基節寬,顎基亦相對向內,顎基刺短,這些特徵形成的應力分布都有助於食殼性能。根據基節形狀的力學模擬,牠甚至能比基節細長的鱟承受更高的負荷。而其特別短而粗壯的顎基刺,也能在咀嚼過程中承受較大的力量。這些強大的基節和顎基刺,顯示霸王萊德利基蟲基節構造特別適合食殼性生活模式。在鴯鶓灣頁岩中,霸王萊德利基蟲與其他受傷的三葉蟲個體以及富含三葉蟲碎片的糞化石共存。以霸王萊德利基蟲的巨大體型(可長達 25 公分)加上其附肢上特有的大型顎基,推測這個種可能就是造成上述受傷三葉蟲和含三葉蟲碎片的糞化石的掠食者,並且可能還會同類相食。
顎基的功能
鋸齒擬油節蟲的附肢形態與上述有所不同。牠有一個卵形到近方形的基節,基節的腹面到顎基內緣都布滿細長像剛毛的顎基刺,這些無疑也能幫助咀嚼功能。力學分析結果顯示,與霸王萊德利基蟲相比,這些剛毛狀刺轉移到基節的應變非常少。細長的顎基刺不太可能承受壓碎硬殼所需的強大力量,這表明鋸齒擬油櫛蟲可能吃軟體食物或腐食,也可能是食碎屑者。這個推論也可以從鱟較前面的步肢的型態與功能得到印證。鱟的附肢 II-IV 的顎基上的棘較長而精緻,無法壓碎貝殼,但可以撕碎較軟的食物,所以鱟用這些附肢咀嚼軟的或預先壓碎的食物,並推動它們進入向後開啟的嘴。鋸齒擬油櫛蟲的卵形基節除了顎基有刺外,在顎基的下方也布滿了短刺和長刺,可能適合向內旋轉以捕獲和切碎軟食物,而不是一個能壓碎帶殼獵物的有效的結構。
軍備競賽
顎基構造廣泛的出現於寒武紀的節肢動物身上,也包括一些非三葉蟲節肢動物。不論其是否能壓碎硬殼,都是一種有效的攝食構造。也許最早演化出來的顎基是為了咀嚼軟質食物,後來特化出能壓碎獵物外殼的結構。似乎在生物演化出礦化外殼的防禦構造後不久,節肢動物便出現了顎基特化為能食殼的掠食種類,上演著獵物與掠食者間永無止息的競賽。


本著作由本館研究人員所提供,博學多文團隊編輯製作,以創用CC 姓名標示–非商業性– 禁止改作 4.0 國際 (CC BY-NC-ND 4.0) 授權條款釋出。若需要使用本篇的文字、圖像等,請洽本館出版室。


