木是植物輸水和支撐的重要構造,泥盆紀的植物僅有初生木質部。石炭紀的鱗木和蘆木,其木質細胞分佈鬆散,稱爲「疏木」。針葉木產生均質紮實的「密木」,適合侏羅紀濕冷的環境,呈現複雜又多樣的「材華」。

生物拓殖陸域空間,離水技能越強的動物夢想飛天,腳踏實地的植物望向天際越長越高,登陸似乎不再是終極目標而已,而是向天演化的歷程。
古生代泥盆紀登陸的植物中,產生維管束的植物在長高、長壯、競爭光線方面,比不具有維管束組織的苔蘚類具優勢,而木質部就是維管束中植物輸水和支撐的重要構造。
早期的木質部
最早的萊尼蕨 (Rhynia sp.) 和庫氏裸蕨 (Cooksonia sp.) 化石,可看出木質部的構造佔很小比例,類似現今的松葉蕨 (Psilotum sp.),木質部很單純,只具有管胞 (tracheid)。古早的蕨類只產生少量初生木質部 (primary xylem),就足以支撐其矮小個體並供應水分。
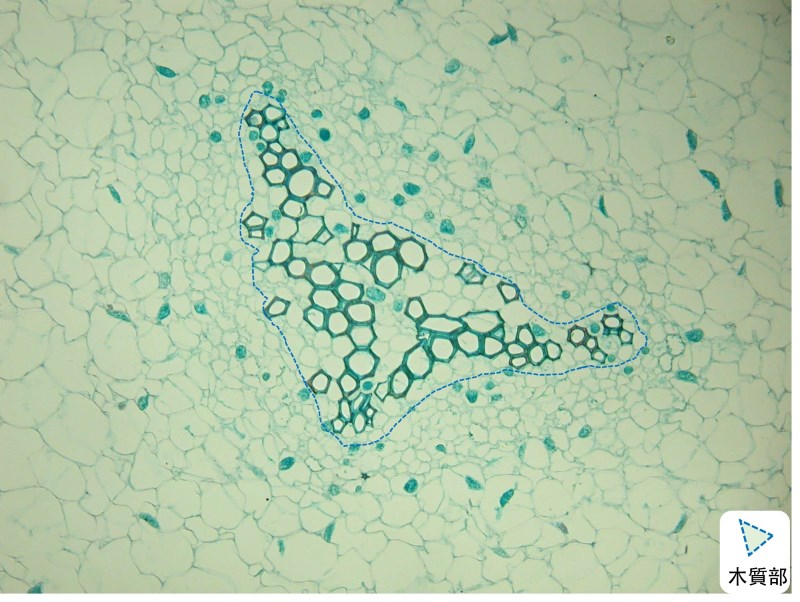
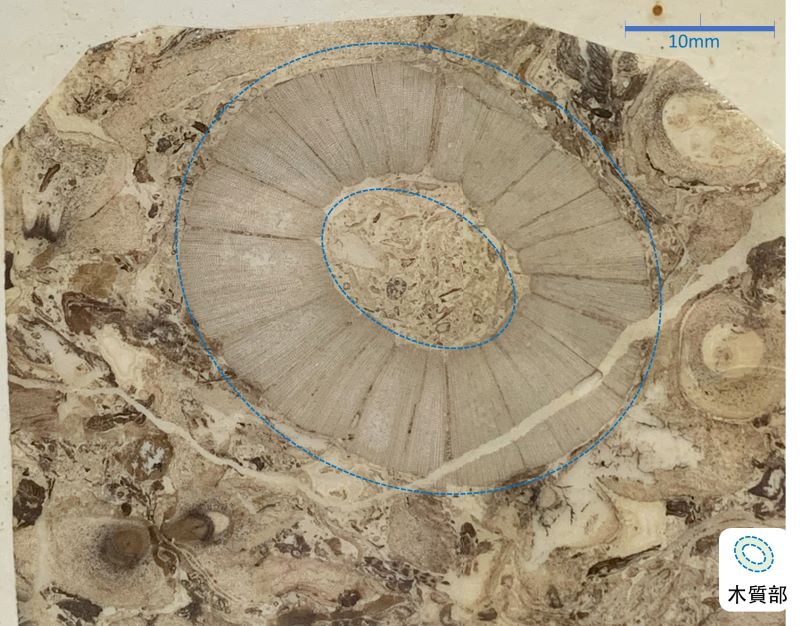
隨著彼此競爭光線,維管束植物越長越高,輸水效率和支撐力需求逐漸提升,在成熟莖內,原本僅分化出生木質部和韌皮部的原始形成層 (procambium),為了增加木質部的量,此時由分化增添具細胞分裂的能力,就是管束形成層 (vascular cambium) 的起源;在演化出木質部長時間增量的過程中,衍生出次生木質部 (secondary xylem) 即木材。
增高增壯的古早蕨類,像放大裸蕨的雙分叉外形,小葉以螺旋狀的序列排列在表皮,掉落後的葉痕讓莖的表面產生類似鱗片的紋理,這些化石具有鱗片狀表面特徵,又產生次生木質部的木材,被歸類在鱗木 (Lepidodendron sp.),是石炭紀地球上最早的森林成員之一。

最古早的森林
雙分叉樹形的鱗木除了壯碩的莖,也衍生出支撐地上部的壯碩根,能連結並支持地上的莖,由直立的莖轉變成固著及具吸收功能的根,同時也產生木材。從地上部轉換到地下部時,外貌由似鱗片狀的葉痕,轉變成連結到主根的圓點痕,就是初生側根的痕跡。由於化石露出大多是片段,幾乎很少是整棵完整的樹,所以這些石頭化的根化石,布滿像肚臍眼的圓點根痕,被歸類到石根 (Stigmaria sp.) 或稱臍根座。


當然古植物學家由更多出土的鱗木、石根化石拼湊建構了初始的鱗木樹樣,根據物種命名規則,以最早發表的學名為正名,完整復原的其他部位名稱就歸為異名,因此石根常常符合了某種鱗木的復原,就變成了異名,而被統稱為鱗木。
最古早的森林中,另有一群以增長節間的方式變高的蕨類,葉排列成一輪,集中在節的輪生,從較矮的楔葉目 (Sphenophyllales),演化至蘆木 (Calamites sp.) 高大樹種,即現生木賊 (Equisetum sp.) 的祖先。石炭紀的森林成形過程中,石松卷柏類與木賊類增高策略的差異,還有植物體形由雙分叉轉化成頂芽優勢的單軸圓錐形,是多樣性森林組成的成功關鍵。

以力學的角度來看,楔葉類植物先整編內部組織結構,空心替代了實心,以空腔管道支撐節間較長、植物體變高的演變;再加上管束形成層產生木材,因此出類拔萃的高大蘆木應運而生。
石炭紀最早的鱗木產生的木材有限,其相鄰的薄壁組織比例仍然很高,而高壯的蘆木則產生較多的木材,空腔組織比例也較鱗木高,因此古早森林的木材本身是較疏鬆的組織結構,因此被稱為「疏木」(manoxylic wood)。另一群源自於種子蕨的樹形蘇鐵類植物,其木材仍屬於疏木型,除此之外,其他的裸子植物則呈現不同的木材緻密度。

更高壯的樹木
不同於髓木 (Medulosa sp.) 和蘇鐵 (Cycas sp.),另一群原始裸子植物中的科達目 (Cordatales),不僅外型更像現今的針葉樹祖先,它的木質部相對於整體組織比例較高,所衍生的木材是較緻密的、均勻一致的管胞,也是地球史上後來出現的銀杏類、松柏類植物的祖先。
科達目的後代歷經氣候變遷,活躍到侏羅紀極盛期,成為最高最壯的森林。最古老生存至今的活化石植物銀杏目 (Ginkgoales),其次生木質部與疏木不同,是一系列排列整齊又緻密均質的管胞,被稱為「密木」(pycnoxylic wood)。僅極少部分穿插輻射向外的射髓 (ray),是髓 (pith) 向外橫面雙向輸送和溝通的管道,大多由薄壁細胞所構成。

隨著侏羅紀末期至白堊紀的環境劇變,被子植物的起源,不僅是開花繁殖構造與傳粉者的共同演化,另一方面也演化出更有效率的輸水構造,也就是導管 (vessel),以因應雨季豐沛水分供給與乾季乾燥缺水間的環境驟變,特別是水澤與沙漠、高山與海岸、極地寒原與赤道熱帶等的極端環境,由管胞和導管細胞不同形態和比例的組合,演化出材華迥異的多樣性闊葉木。
裸子植物中的蘇鐵源自於種子蕨,它的木材仍屬於疏木型,樹型長高有限。而科達目的後代因應生存競爭及氣候變遷,活躍於侏羅紀成為最高最壯的極盛森林。從適應侏羅紀的濕冷環境,活化石的銀杏和針葉木才有較大比例、密實木質化管細胞的木材,相對被稱為「密木」。
歷經地球上氣候變遷及環境驟變,白堊紀才發展出具導管細胞的闊葉木,是組合複雜多樣的木材,與針葉木同屬密木型。適應與演化產生橋木、灌木、木質藤本多形多樣生活型,也使木材呈現出各自出眾的「材華」。
後記:
石炭紀時的鱗木和蘆木,木質細胞鬆散、不紮實,產生木材量不多,反而內含較多的薄壁細胞。從化石撕片和磨片中可以看見微小疏鬆的古早木材,屬於疏木型。而侏羅紀高大森林是現今密木型的銀杏、松柏的祖先。
1. 鱗木化石 (Lepidodendron sp.)
已滅絕的石松卷柏類樹狀祖先,樹幹表面具密集的菱形葉座、似魚鱗狀的樹皮。
2.鱗木/石根 (Lepidodendron / Stigmaria sp.)
樹狀石松植物的地下根結構。
3.臍根座 (Stigmaria ficoides) 鱗木類樹幹的基部,表面有臍狀根痕,為不定根著生的地方。
4.楔葉科化石 (Sphenophyllaceae)
已滅絕的木賊類,莖枝細弱,分節與節間,節長輪生葉。
5.蘆木化石 (Calamite sp.)
已滅絕的木賊類高大的樹狀祖先,與鱗木、封印木等蕨類形成最早的石炭紀森林。
6.蘇鐵化石(Cycas sp.)
種子植物最古早的活化石,適應暖熱的氣候,是古氣候和古地理研究的重要指標。
7.銀杏 (Ginkgo biloba)
壽命可達千年以上的落葉喬木,最早的密木活化石,可追溯到 2.7 億年前。
8.中新世種子植物木化石

延伸閱讀

本著作由本館研究人員所提供,博學多文團隊編輯製作,以創用CC 姓名標示–非商業性– 禁止改作 4.0 國際 (CC BY-NC-ND 4.0) 授權條款釋出。若需要使用本篇的文字、圖像等,請洽本館出版室。


