

上世紀後半到本世紀在非洲與亞洲出土的恐龍化石和新發現,大大填補了中到晚侏羅世間的演化空缺,從而解釋了晚侏羅世恐龍的高多樣性由來。當中最重要的發現之一,是找到了羽毛的起源,以及從龍到鳥最初的2千萬年的歷史。

晚侏羅世公園
在整個侏羅紀的5600-6000萬年中,後1/3的晚侏羅世(1.63-1.45億年前,起訖尚有爭議)是真正符合流行文化想像的《侏羅紀公園》:大眾熟悉的迷惑龍(Apatosaurus)、腕龍(Brachiosaurus)、圓頂龍(Camarasaurus)、梁龍(Diplodocus)等巨大的植食獸、異特龍(Allosaurus)與角鼻龍(Ceratosaurus)等大型掠食者、小巧靈活的美頷龍(Compsognathus)、長著骨板並帶有尾刺禦敵的劍龍(Stegosaurus)及有著直拇指的彎龍(Camptosaurus)等都已登場。
除了《侏羅紀》系列電影的影響,這些印象最早是源自19世紀後半的〈骨頭大戰〉(Bone Wars):兩位美國的恐龍研究者使盡各種手段在西部競逐化石,阻撓甚至破壞對手的挖掘,並相互詆毀攻訐。最終雙方散盡家財也賠上名聲,至死方休。兩人共命名了136種恐龍,看似成績斐然,但他們常草率地將零散無法鑑別的化石命名為種;或不管對手的發現,兀自命名手頭上的類似材料,導致同物異名;甚至將非同一處挖到的骨頭拼湊成種,造成混亂。經過後人重新檢驗比對,現在被承認的只剩32種,但多半是大眾耳熟能詳的物種。

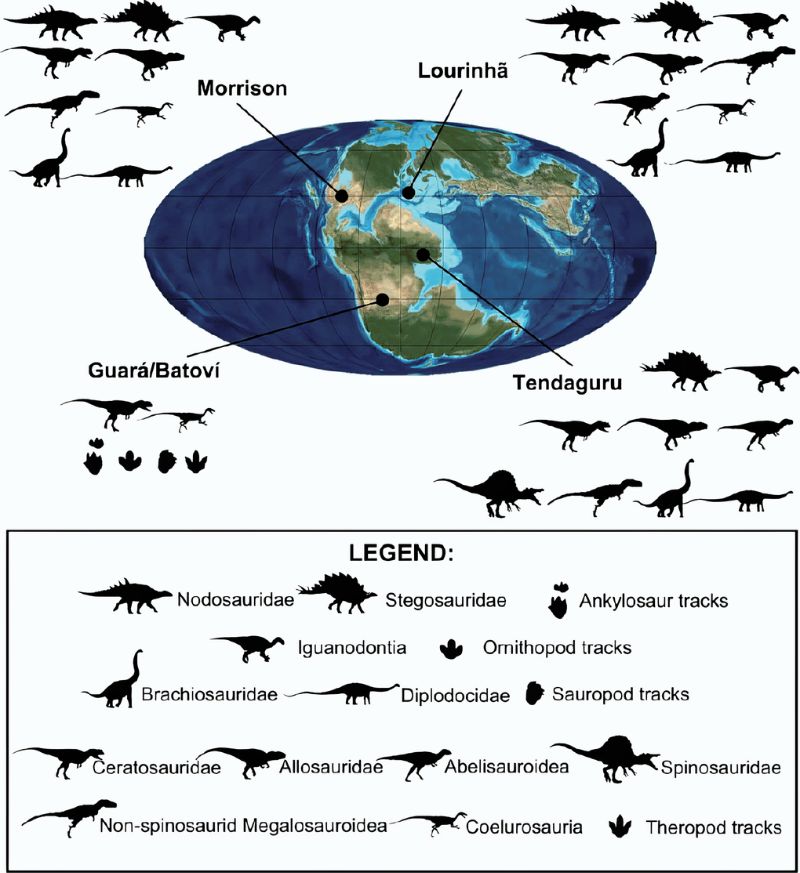
另一方面,這段歷史也讓北美從原本只有9種恐龍的紀錄在20多年間暴增至百餘種,使得古生物研究前線從西歐轉移到美國,引發之後超過一世紀的恐龍研究熱潮,至今不退,並將恐龍打進大眾文化。〈骨頭大戰〉的一個遺緒,是1877年在莫里遜層(Morrison Formation,屬啟莫里~提通階,1.57-1.47億年前)發現大量且多樣性極高的恐龍,也是雙方角力的熱點。範圍廣達數州的莫里遜層至今仍是美國恐龍化石最豐富的地層,累計達37個屬,是經典的晚侏羅世恐龍相。連載14會再回頭談這兒的故事。

在世界其他地區,如葡萄牙的洛里尼彥層(Lourinhã Formation)、東非坦尚尼亞的天達古乎層(Tendaguru Formation)等與莫里遜層約同時期的地層,也都發現多樣性極高的恐龍。雖然美西的莫里遜層主要為洪積平原(alluvial plains),洛里尼彥層與天達古乎層則是海岸地形(有海相化石),但彼此的恐龍相相似性頗高。這說明盤古大陸雖已裂解為南北兩大陸,北方的勞亞大陸也分離為東西兩大陸塊,中間散布著大小不一的島嶼(歐洲),但大陸與陸塊間仍存在著廊道,能互通有無。
蓄勢待發的黎明
跟晦暗不明的中侏羅世相比,晚侏羅世令人驚奇的恐龍多樣性猶如從天而降,來歷(演化脈絡)不明。其實晚侏羅世最初的400-600萬年(牛津期),早期記錄到的恐龍多樣性同樣乏善可陳。但上世紀後半到本世紀,在非洲和亞洲的挖掘與研究獲得許多突破性的進展。例如在中國四川、重慶、內蒙、準噶爾盆地等都發現不少從中侏羅世晚期到晚侏羅世初期的恐龍。當中不乏殘存的舊支系和新支系的初始物種,填補許多形態、多樣性變化與譜系的空缺。
以四川盆地的下/上沙溪廟層為例,至今發現超過30屬各類恐龍,當中的渝洲龍(Yuzhoulong)與大山鋪龍(Dashanpusaurus)是最早的大鼻龍類(macronarians,稍晚的圓頂龍、腕龍等皆屬之),也顯示牠們在中侏羅世便已擴及全世界;準噶爾盆地的石樹溝層累計達17屬,有亞洲最早的角鼻龍類泥潭龍(Limusaurus)、暴龍總科(tyrannosauroids)中極初期的冠龍(Guanlong),以及最早的角龍類(ceratopsians)隱龍(Yinlong);內蒙寧夏的直羅層出土的靈武龍(Lingwulong)是梁龍總科(diplodocoids)唯一的亞洲物種;燕遼生物群(Yanliao Biota,包含內蒙海房溝層的道虎溝生物群,和冀北/遼西髫髻山層的玲瓏塔生物群)也出土11屬恐龍,除鳥臀目畸齒龍科殘存的天宇龍(Tianyulong),其餘多半是跟「鳥類」相關的小型獸腳類,與早白堊世著名的熱河生物群(Jehol Biota)相關聯。


不過這些地層和定年尚有爭議。例如靈武龍原以為是出自中侏羅系巴柔階的延安層,後來認定是來自其上層的直羅層,屬巴通~牛津階;在不同地點對下沙溪廟層底部鈾鉛定年,出現1.69、1.66和1.61億年前的差異,有可能是強烈侵蝕導致跨時(diachronous)沉積,或是地層認定的不確定性;此外,石樹溝層跨越約1千萬年(1.65-1.55億年前);道虎溝與玲瓏塔生物群分別約1.67與1.61億年前。由於此間的恐龍呈現相當的連續性,因此多半以中~晚侏羅世歸屬之。
從龍到鳥之路
在演化史上,這段期間還發生一件大事,就是「鳥類」的出現。1860年代初在德國發現的始祖鳥化石兼具恐龍與飛鳥特徵,似乎印證了達爾文物種原始中預期的過渡型式,也就是所謂失落環節(missing link)的存在,而在「達爾文的牛頭犬」赫胥黎(Thomas H. Huxley, 1825-1895)的闡釋下,開啟了龍/鳥關係將近150年來的辯證。學界現在普遍接受〈鳥即恐龍說〉,這主要歸功於1990年代中期起一連串對中~晚侏羅世的新發現。
由於始祖鳥給人印記般的印象,上世紀有很長一段時間將羽毛等同飛行,也當成是鳥和恐龍的分界。但隨著熱河與燕遼生物群中許多似鳥類龍的生物出土,顛覆了此一界定。羽毛不僅有著漸進的演化過程,且不侷限於獸腳類與鳥;飛行也非「鳥」專屬,在「非鳥恐龍」有過許多成敗不一的騰空嘗試。
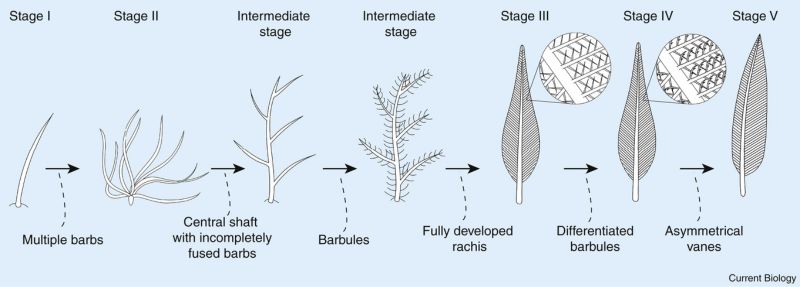
2014年發表了中侏羅世巴通期(1.68億年前)的一種初期新鳥臀類恐龍:庫林達奔龍(Kulindadromeus),同時帶有鱗片和羽毛。2009年發表自髫髻山層(1.59億年前)的天宇龍也帶有初期羽毛。兩者都屬鳥臀類,與獸腳類關係很遠,卻都有羽毛,而3大類恐龍中只有蜥腳形類尚無證據。學者對此有不同解釋:1)羽毛可能是恐龍的祖徵,之後多次丟失;2)不同類群都有長羽毛的遺傳潛力;3)獸腳類與鳥臀類是姊妹群(請參考:恐龍便便裡的名堂(10))。這有賴發現更早的羽毛來驗證。

獸腳類的羽毛可從鳥翼類(包含所有現生的鳥)回溯至堅尾龍類。除少數例外,各主要分支內皆出現不同演化階段的羽毛。美頷龍起的手盜龍形類開始有小型化靈巧的趨勢;「鳥翅」般的前肢出現在手盜龍型類;廓羽盜龍類始有「正羽」(pennaceous/contour feather,=「廓羽」);在近鳥類中,鳥翼類以外的其他小型物種多半鳥模鳥樣,稍大型物種如馳龍科(Dromaeosauridae)的振元龍(Zhenyuanlong)翅上有正羽,知名的恐爪龍(Deinonychus)雖然尚未發現羽毛跡證,但推測也是帶羽恐龍。
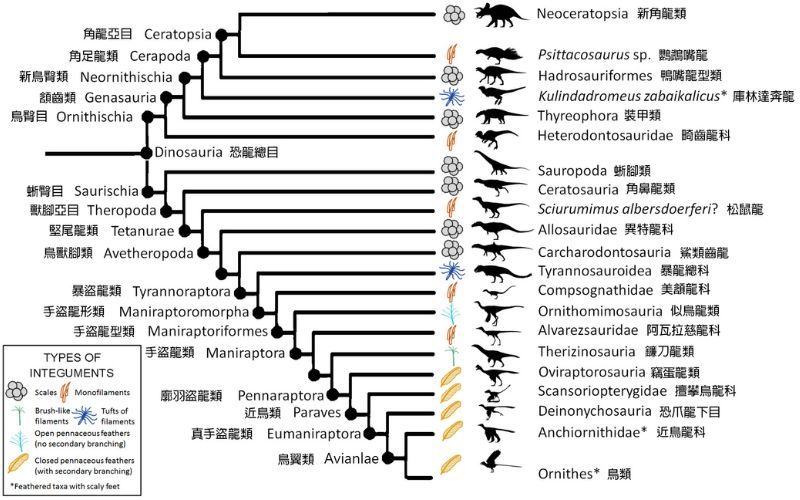

至於飛行能力,直觀上始於鳥翼類祖先,但也有不少學者主張起源於更早的近鳥類甚至廓羽盜龍類祖先,只不過後來多數地棲類群失去此能力,僅鳥翼類保留下來;還有學者認為從晚侏羅世到整個白堊紀,飛行在近鳥類各支系中獨立發生數回,但只有鳥翼類存續下來,演化成現在的鳥。
當鳥不再是鳥
上世紀末以來的許多新發現,讓恐龍與鳥類之間不再涇渭分明,也讓鳥的演化上溯千萬年。始祖鳥在不同分析中被歸為鳥翼類或真手盜龍類,是否能稱之為鳥已非牢不可破。連帶地,牠不再是標誌般存在的「鳥」始祖,也非龍鳥之間神奇的失落環節,而是連環套中的一環。什麼是「鳥」取決於譜系節點上何者為鳥的認定,比較像語意學的問題,而非本質上的差異。
化石紀錄上,與「鳥」最相關的恐龍中(近鳥類),以燕遼生物群的近鳥龍科(Anchiornithidae)與擅攀鳥龍科(Scansoriopterygidae)最早。雖然牠們的大眾知名度遠不如始祖鳥,但其科學重要性卻不下於後者,對鳥的定義與飛行起緣理論帶來極大衝擊。牠們比始祖鳥早了至少1700萬年,解決了1990年代後期所提出的(始祖)鳥早於(小盜)龍的時間悖論(temporal paradox)。牠們有羽有翅,可能也是最早能騰空的恐龍,讓龍鳥之分更加模糊,也更加支持鳥就是龍的假說。我們會在下篇連載作更多介紹。
正模式中的正模式
最後談一下這段歷史中的食渣化石。連載11提到在波蘭的恐龍糞化石中發現新種化石植物,而恐龍食渣化石中也發現過其他新種化石動物。
直到1990年代之前,美頷龍(Compognathus)都是最小的非鳥恐龍紀錄保持者,體長連尾巴僅70多公分,肩高只有20公分左右。牠們是少數出現在侏羅紀的鳥的遠親,所在地層與始祖鳥相同或相近,屬上侏羅統提通階,距今約1.5億年前。沉積環境與化石顯示當時當地為海岸潟湖,共棲的除了始祖鳥,還有一些翼龍,但無大型恐龍。同地層還出土許多海洋脊椎與無脊椎動物。
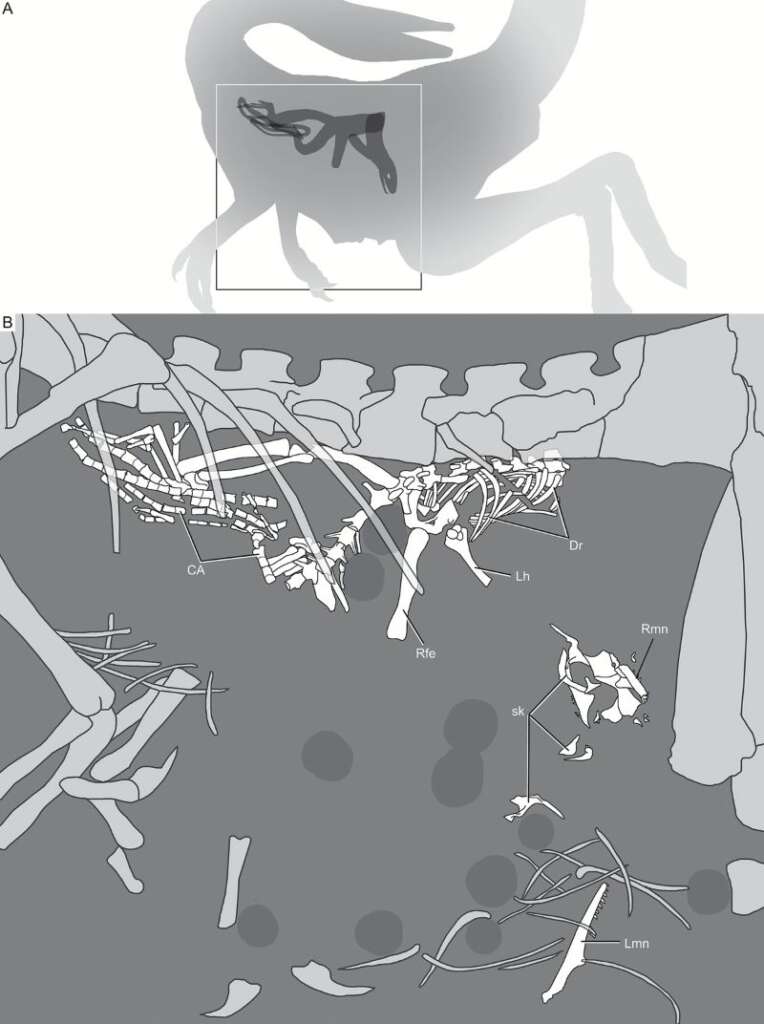
德國出土的長肢美頷龍的正模式標本相當完整,骨架大半都還互相連接,顯示死後不久或溺斃後很快便沉入海中被掩埋,未被動物分食或細菌分解。其體內很早便發現含有其他動物骨頭。馬許在1881年檢查該件標本後,認為殘骨是留在體內的胚胎(換句話說他認為美頷龍是胎生);之後的學者則鑑定出是一隻蜥蜴,可能為巴伐利亞蜥(Bavarisaurus)。由於蜥蜴基本完整,許多關節仍相連結而非散骨,應是整隻吞下而非食腐攝入。 2017年學者以該件胃含物為正模式標本,命名為新屬新種蜥蜴Schoenesmahl dyspepsia,成為正模式標本含有正模式標本的奇特案例。這也顯示美頷龍是靈巧迅速的獵食者,能捕食快速活動的獵物。


延伸閱讀

本著作由本館研究人員所提供,博學多文團隊編輯製作,以創用CC 姓名標示–非商業性– 禁止改作 4.0 國際 (CC BY-NC-ND 4.0) 授權條款釋出。若需要使用本篇的文字、圖像等,請洽本館出版室。


