
翼龍在 2.1 億年前的晚三疊世就飛上了天。但牠們的親戚,也就是恐龍,要到 1.6 億多年前的中侏羅世才開始飛行的嘗試。在最初 2 千萬年裡,牠們做了不同的嘗試,雖有羽翼且鳥模鳥樣,但離真正能飛還有很長的路要走。本篇就來談談從龍到鳥在侏羅紀的極初期演化。

霓裳羽翼初長成
先談近鳥龍科。本科原被歸於恐爪龍下目的傷齒龍科 (Troodontidae),2017 年才獨立出來(非所有學者都認同),成員也都在本世紀才被發現或命名。現含 9 屬中,除歐斯壯龍(Ostromia,2017 年歸入本科)產於德國,餘皆源自中國;最早的足羽龍 (Pedopenna) 屬道虎溝生物群(1.67 億年前),多數為玲瓏塔生物群(1.61 億年前),歐斯壯龍屬晚侏羅世後期(1.5 億年前);義縣龍 (Yixianosaurus) 則屬早白堊世的熱河生物群(1.31 億年前)。另有遼寧獵龍 (Liaoningvenator) 是潛在成員,但有學者認為牠與義縣龍(或全部 10 屬)是傷齒龍科。
本科對鳥與飛行的早期演化提供了眾多資訊。例如羽足龍雖然只發現一個後肢,但上頭帶著整排正羽(IV 級),較晚的近鳥龍 (Anchiornis)、絲鳥龍 (Serikornis)、彩虹龍 (Caihong) 和曉廷龍 (Xiaotingia) 也是,跟白堊紀的小盜龍類 (microraptorians) 有相仿的四翅外觀,卻不見於始祖鳥。這支持「四翅」是近鳥類或真手盜龍類的祖徵。但小盜龍後肢帶飛羽(V 級),是名符其實的「後翅」,在近鳥龍科則尚無氣動性 (aerodynamic)。這也說明後肢羽列最初無涉飛行。

2018 年發表的彩虹龍,在頭頸部、胸部和尾巴基部的羽毛帶有類似現生蜂鳥餅狀排列的黑色素體 (melanosomes),很可能有斑斕的色彩;牠的前肢骨比其他近鳥龍短,但帶有更長的正羽來增加翅面積;芭蕉扇般的尾比始祖鳥更大,而且帶 V 級羽,是迄今最古老的不對稱羽化石。這也意味飛羽最初是由尾巴發展,而非前肢,或許尾巴在飛行起源上有某些功能。

而令人好奇的是近鳥龍科雖然「鳥模鳥樣」,但牠們能飛嗎?跟始祖鳥相比又如何呢?
攀上枝頭之後
擅攀鳥龍科是龍鳥之路上的怪客。牠們也是在本世紀才被發現與建立,目前有 4 個成員:擅攀鳥龍(Scansoriopteryx,樹息龍 Epidendrosaurus 為同物異名)、耀龍 (Epidexipteryx) 與渾元龍 (Ambopteryx) 屬道虎溝生物群,奇翼龍 (Yi) 則為玲瓏塔生物群。擅攀鳥龍與耀龍所根據的標本都是幼獸/雛,並未引起太多討論,直到 2015 與 2019 年以成體或亞成體標本發表的奇翼龍與渾元龍相繼問世,擅攀鳥龍科的奇異之處才引發熱議。

跟其他近鳥類恐龍相較,牠們長得很特異:1. 吻部短;2. 前肢比後肢長、尺骨比掌骨長、三指中底下一指最長、腕骨特化出針狀骨結構 (styliform element);3. 尾椎短且癒合為尾綜骨 (pygostyle),與現今鳥類相仿,始祖鳥反倒有未癒合的長尾椎;4. 有兩對裝飾性尾羽;5. 後肢有羽列,前肢前緣有 II – III 級羽,後部卻帶有翼膜!這些特徵中,針狀骨與翼膜未曾在其他鳥類出現過,尾綜骨則是首度出現。

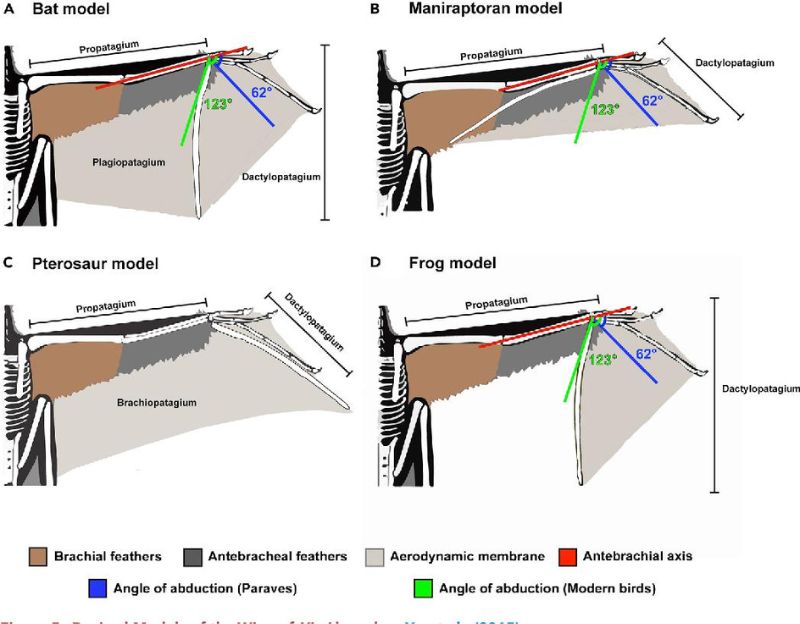
樹棲滑翔哺乳類如鼯鼠也有針狀骨,可以撐大翼膜。或許擅攀鳥龍科也具有類似構造,用長指爬上樹後,再一躍而下以翼膜騰空。然而翼龍與蝙蝠靠翼膜動力飛行,哺乳類與爬蟲則藉以滑翔,那麼擅攀鳥龍科呢?
想飛?難如登天!
侏羅紀這些龍/鳥先驅者能飛與否,直接牽涉飛行起源的大哉問。相關研究汗牛充棟,且仍在進行。研究邏輯不外與現生鳥類飛行所需條件比較,例如體型大小、有無飛羽、翅的構造與面積、飛行肌肉與骨架、胸腔與心臟強度……看這些半龍半鳥動物(特別是始祖鳥)的符合度,找出現生鳥類中接近的類比。
既然如此,就得先了解鳥類飛行骨架-肌肉的基本構造與運作。飛鳥的肩胛骨、鳥喙骨和叉骨構成適於飛行的肩帶,肩臼(肩窩)朝側上方,讓肱骨頭有較大的活動範圍;叉骨能維持振翅時胸廓的穩定又保有彈性;鳥喙骨下接胸骨,控制飛行的兩條主要肌肉著生在胸骨的龍骨突起上;喙上肌肌腱穿過三骨管繞接到肱骨頭背面,收縮時能像滑輪組一樣讓翅上舉;厚實的胸大肌則連接肱骨頭腹面,負責翅的下壓,藉振復輪迴產生昇力與推力動力飛行。
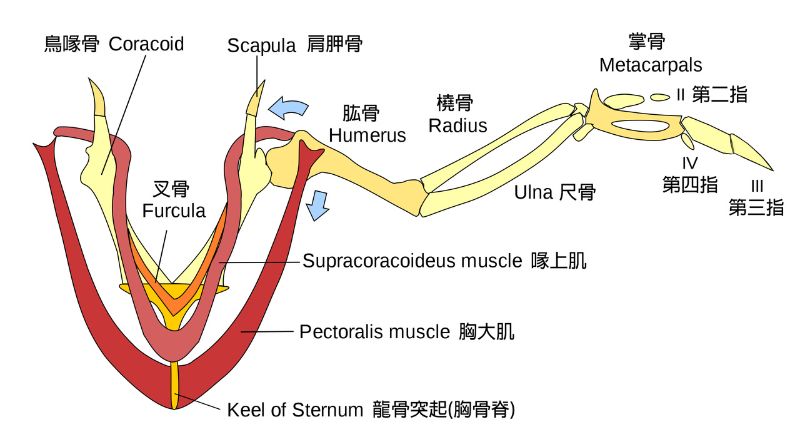
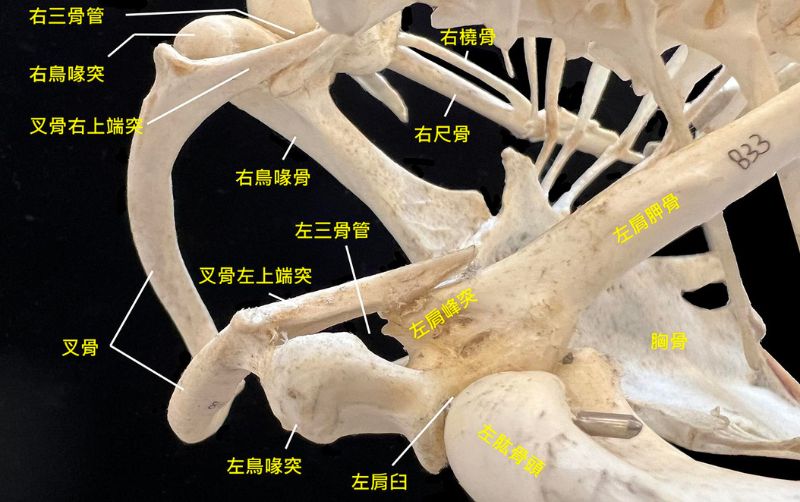
不過鳥類有各種飛行樣式,如疾振翅懸停、靠熱氣流盤旋、遠距離滑翔、短距離快飛等,還有許多失去飛行能力的鳥,其飛行骨架與肌肉都有所差異。2023 年的一篇報告分析 600 多種現生鳥類胸大肌與喙上肌的質量比,發現與許多行為生態差異相關。例如胸大肌隨體型大小增減,喙上肌則相反;能懸停的蜂鳥有不成比例的強大喙上肌,短距離快飛的鳥也不小,猛禽則相反,肌肉量比甚至達 20 : 1。不過構造萬變不離其宗,在此不加細述。
演化史上能飛的脊椎動物類群屈指可數,成功是罕見事件,「想飛」真的是「難如登天」!蓋飛行是系統性運動,需要結構間緊密搭配。單從表相或單點切入雖能測試一些基本條件(如翼負荷、昇力、速度等),但未必可知到底能不能飛。以下僅就本人認為較有系統觀且具說服力的近期研究作介紹。
龍/鳥最初的兩千萬年
侏羅紀的龍/鳥先驅尚未發展出適應飛行的骨架,因此肌肉著生位置與作用(化石看不到)必然不同。直接拿現生鳥類的構造與機制類比未必恰當,反而必須從系列化石中找出脈絡,去理解這些結構與運作方式的改變對飛行的影響。
首先,這些龍/鳥先驅不僅沒有較發達的胸骨,反而完全丟失,代之以腹肋骨來支撐胸腔。這也許有助於呼吸換氣,但無法提供強大肌肉附著,化石的腹部輪廓也反映近鳥龍的胸大肌並不發達。喙上肌則著生在鳥喙骨板上,不會像現生鳥類長在龍骨突起上那般發達。其次,龍/鳥先驅者的肩帶雖然已從較腹面位置向前上方位移,且呈現夾角,但尚在身體兩側,構造也較原始。其肩臼朝向側面,限制了肱骨上舉的角度。
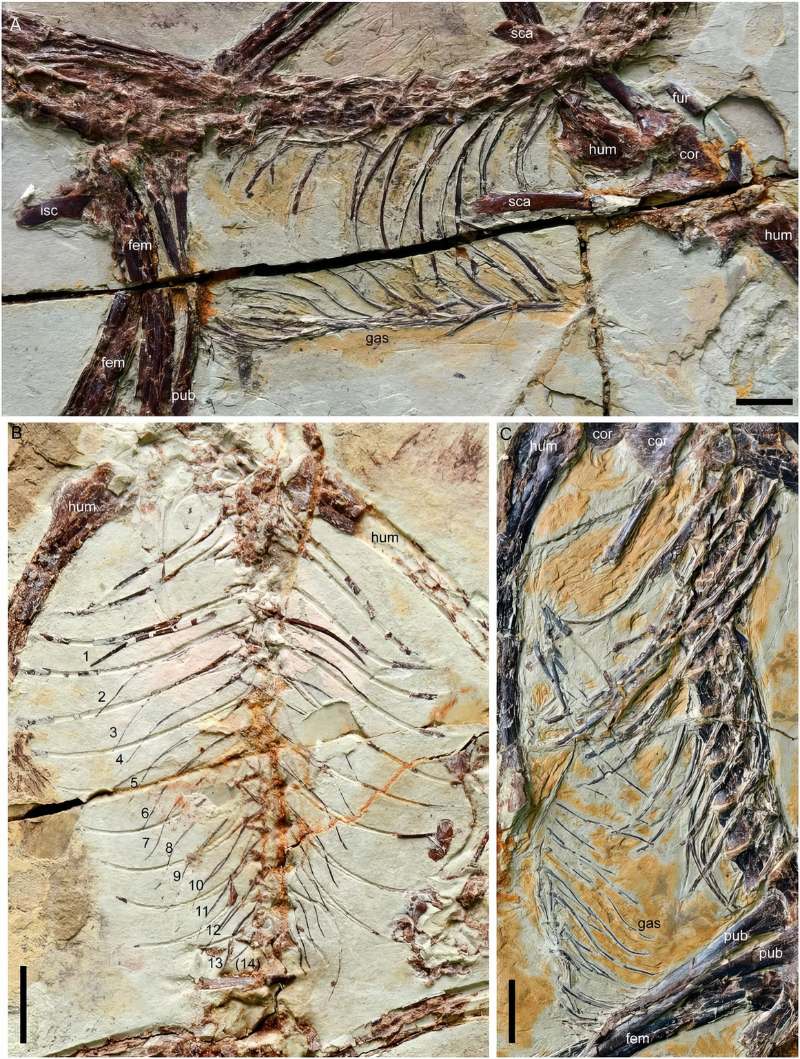
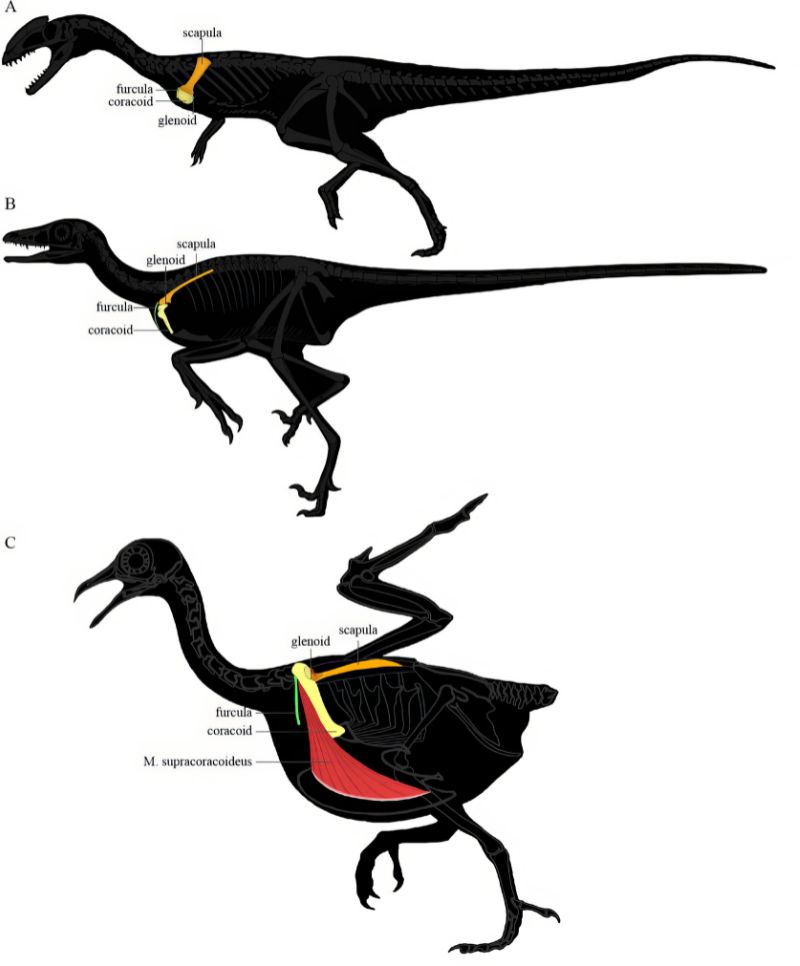
而隨著肩帶位移與形變,原本負責前肢後收的喙上肌功能也跟著轉變。始祖鳥與近鳥龍的鳥喙突比同時代的獸腳類發達,但仍低於肩臼,尚未形成三骨管;喙上肌的肌腱繞過鳥喙突後只達肩臼前端,著生在肱骨頭前側,因此收縮時牽動的方向是朝前而非朝上。缺少喙上肌的參與,若要上下振翅主要得靠背上的三角肌 (deltoids) 搭配胸大肌,但兩者都不發達。可以想見牠們很難維持振翅的循環,即使有動力飛行能力也應該很有限。
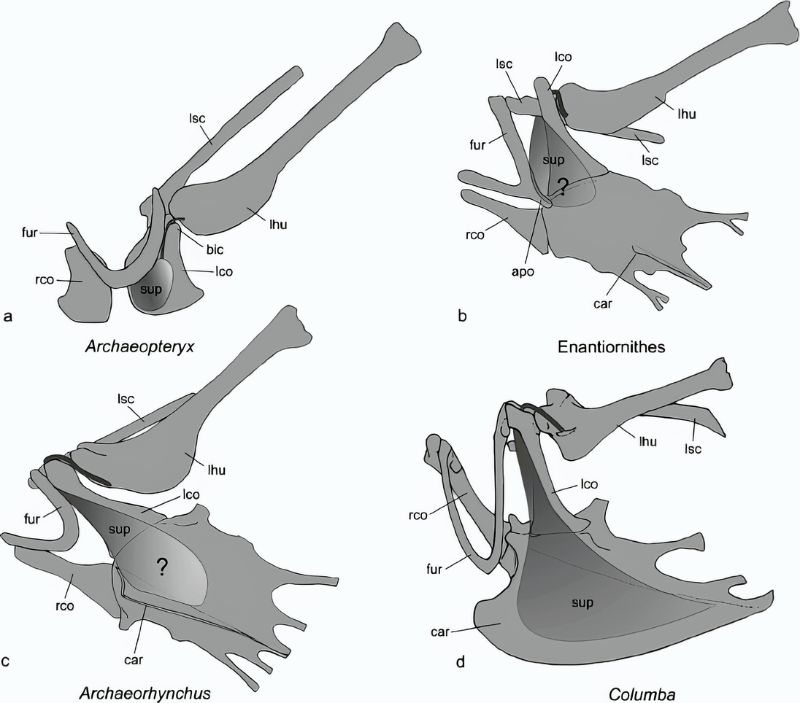
以整體結構跟機制來說,始祖鳥與近鳥龍科反而跟鴕鳥、鴯鶓、鶴鴕等不會飛的古顎鳥(palaeognaths = ratites,平胸鳥)類似,不過牠們身體小得多,輕量的骨頭類似飛禽,而非厚重的古顎鳥。或許能藉翅膀的幫助快跑衝刺,之後短距離騰空,或是滑翔,或跳躍來跨越障礙。但這個階段的羽和翅顯然還不是用來飛行,更可能是展示、威嚇、孵卵、疾奔時保持平衡與快速轉向、能「伸手」攔截捕捉快跑或高處的獵物,或快逃躲避天敵等。以後見之明看,這些功能是成功的,有持續發展下去,最終在千萬年後飛上青天。

至於擅攀鳥龍科,2016 – 2020 年的研究同樣發現牠們的身體、前肢和翼膜結構都限制其拍翅,也不適合疾走,不可能從地面振翅或靠助跑起飛,但或許能藉翅膜在枝幹間上下躍動。至於滑翔,則取決於翼膜構形。雖然無論大小都有些滯空能力(最低如降落傘模式),但唯有翼膜涵蓋前後肢,讓升力點 (center of lift) 後移至身體重心稍後位置,才能滿足滑翔的條件。因此,若擅攀鳥龍科能滑翔,會像鼯鼠以四肢撐開翼膜的模樣。2019 年渾元龍發表時,原作者認為翼膜是早期的飛行嘗試之一,但並不怎麼成功,未能擴散或流傳下來。
直到侏羅紀結束,近鳥類只在東亞出現高多樣性,其他地區則很零星,如歐洲的始祖鳥和從牠分出來的歐斯壯龍(「哈勒姆標本」)、維爾恩胡芬鳥(Wellnhoferia,「索爾恩霍芬標本」)、阿克摩納鳥(Alcmonavis,「第 13 號標本」),與美國的西鳥形龍(Hesperornithoides,傷齒龍科)等,全都位於勞亞大陸。要再過 2,500 萬年,趨近現代飛鳥的飛行結構才逐步演化並擴散開來。
龍/鳥們的產地與食物
從燕遼生物群化石與沉積跡證推測,當時的古環境應是溫帶山區的潮濕森林,環繞著湖泊或湖泊環繞山區,附近有火山活動。當中棲息著多樣的陸棲與淡水棲的無脊椎與中小型脊椎動物,包含許多中小型翼龍,但無大型恐龍。此地層也發現最早能滑翔的哺乳類,如海房溝層的翔獸 (Volaticotherium),髫髻山層的翔齒獸 (Vilevolodon) 與祖翼獸 (Maiopatagium) 等。另一方面,始祖鳥是住在亞熱帶淺海環繞的島嶼,同樣有翼龍卻無大型恐龍。島嶼在演化上是產生新特徵 (novelty) 的重要場域。或許侏羅紀這些島嶼以及如島嶼般的森林提供了飛行演化的驅力與實驗場,讓動物們去發展動力飛行或滑翔,並產生選汰。
最後談談侏羅紀龍/鳥先驅者的食物。始祖鳥雖然發現最久,但迄今僅有十來件標本,未發現任何食渣化石。一份 2021 年的報告提到牠們牙齒的微磨損狀態與現生食蟲性鳥類最接近,不過因為樣本數很少,尚難對食性下定論。
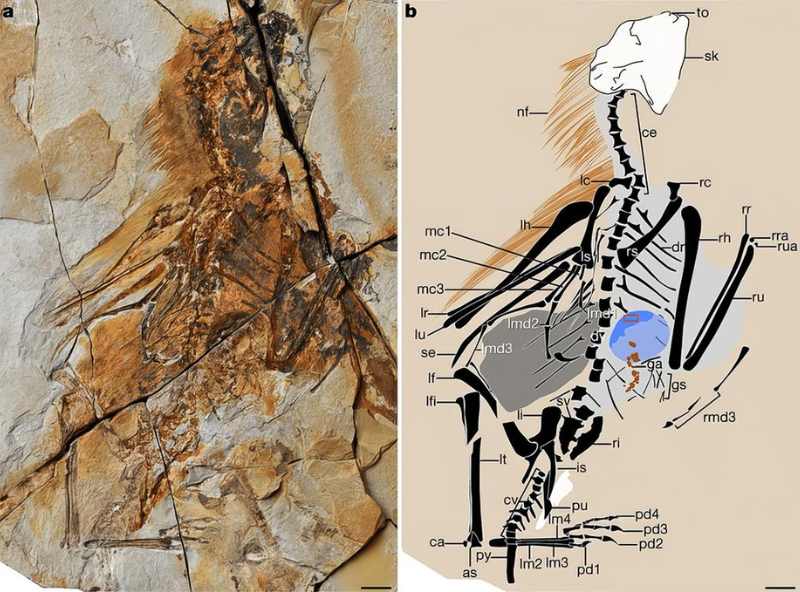
赫氏近鳥龍出土的標本已有數百件,因此得以重建許多生態習性。2018 年的一個研究找到 4 件含有食渣的標本,其中 1 件在食道部位有團狀物,3 件落在骨架旁,另在化石床找到 2 件獨立的橢圓狀物。食道的團狀物內含至少 3 隻蜥蜴的散骨,因此不是剛吞入的食物,而極可能是無法消化、正要吐出的食繭 (pellets)。骨架旁的團狀物形態與此類似,至少有 1 件包含魚鱗。食繭大小跟骨架尺寸呈正相關,應是不同年齡個體所產出。


獨立橢圓狀物則飽含魚鱗與魚骨,應為食繭無誤。若是糞化石,很難會有這麼多鱗片與骨頭能通過消化道保留下來。不過當地還有眾多體型相仿的動物,如擅攀龍和小型翼龍,也是潛在的產出者。但至少食道內的食繭確證近鳥龍是敏捷的獵人,能捕食小型蜥蜴這類快速移動的動物,也許羽翅在追獵時有幫上忙。
擅攀鳥龍科的渾元龍的正模式標本體內也有食渣痕跡,可能是小型脊椎動物殘骨。此外還有疑似胃石 (gastrolites)。因此渾元龍可能是雜食性,獵取昆蟲及小型脊椎動物,並在樹上或下地覓食植物,靠胃石協助消化。(待續)


本著作由本館研究人員所提供,博學多文團隊編輯製作,以創用CC 姓名標示–非商業性– 禁止改作 4.0 國際 (CC BY-NC-ND 4.0) 授權條款釋出。若需要使用本篇的文字、圖像等,請洽本館出版室。


