圖/廖鎮磐提供

風干擾可誘發結網蜘蛛的大壺狀腺絲在機械性質上的可塑性,此種可塑性乃基於絲微觀結構的排列方式不同所致,推論蜘蛛在經歷風干擾後可主動改變體內生產絲之微環境,即使在原料絲蛋白相同的條件下,可快速地改變絲生產過程進而增強絲強度與韌度。

蜘蛛世界的風干擾與絲質適應:捕食者的演化之謎
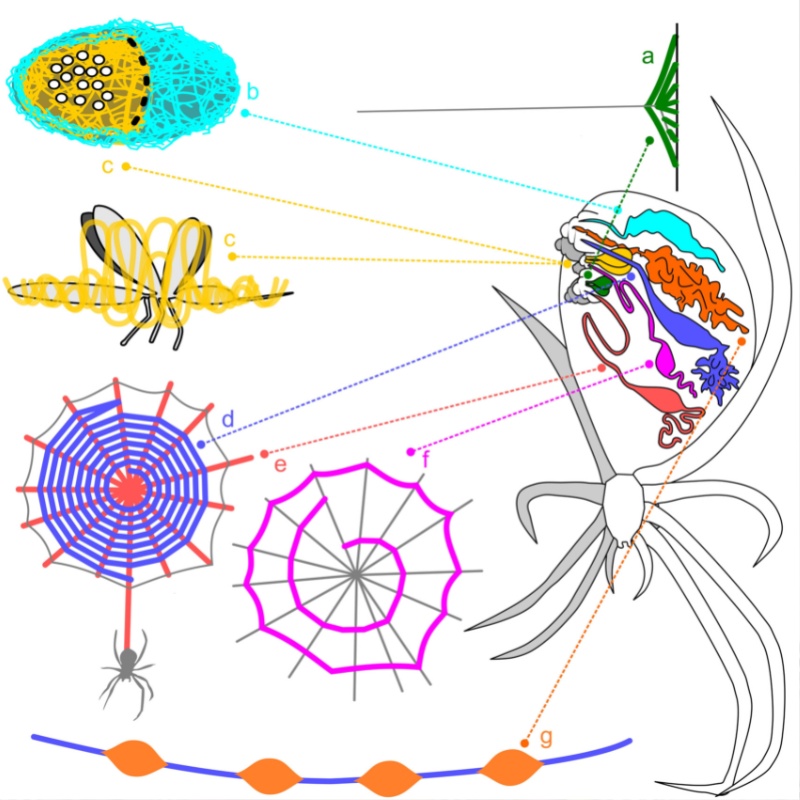
在動物界中,以自身分泌物製作陷阱以捕食獵物的捕食者以蜘蛛最為常見。蜘蛛約在 2 億年前由較原始的地表巢穴伏擊的方式演化至在地表上方的三維空間中建構現今常見的圓網,以攔截大量在空間中飛行的無脊椎動物 (Blackledge, 2009),而成為現今最主要的飛行性昆蟲捕食者 (Vollrath and Selden, 2007)。在其由地表至空中的演化過程之中,除了在捕食行為上表現出差異之外,蜘蛛絲分泌系統也朝向多樣與分工的方向演化。在現今常見的結網蜘蛛中可以同時發現7種具有不同功能的蜘蛛絲,其分泌之腺體、蜘蛛絲蛋白與基因及分泌過程所經過之絲疣皆已完全分化 (Foelix, 2011)。由於蜘蛛網的主要功能是在空中攔截獵物,過程中必須承受獵物的飛行動能再將其轉換成動能。這項功能主要是由大壺狀線絲所構成的輻射狀絲,見下圖 e。在拉伸再收縮的過程中達成,因此可以猜測大壺狀線絲的特性主要朝向高強度(斷裂前的最大張力)、高韌度(斷裂前的最大動能)、適中的楊氏係數(絲張力與延長量的比值)與延展性(斷裂前的延長量)的方向特化。
在由地表演化至到空中的過程中,蜘蛛與蜘蛛網也面對了最主要的一項非生物性干擾因子—風。在行為學上,風干擾增加蜘蛛處理獵物所需時間及獵物由網上掙脫的機率 (Turner et al., 2011) ;風干擾造成相鄰黏絲交纏而造成網目增加;被風完全破壞的蜘蛛網等同蜘蛛浪費了牠結網所消耗的能量與時間並損失所有可能捕捉獵物的機會。綜上所述,風干擾在不同層次對蜘蛛捕食獵物之能量投資效率比皆呈嚴重的負向影響。就此而言,結網蜘蛛的結網行為或蜘蛛絲性質應具有特定適應或可塑性以降低風干擾之影響。先前研究已明確指出,在風干擾盛行之處,蜘蛛可以藉由將網的展開方向與風向平行 (Ramirez, 2003) 、或減少網面積或增加網目寬度 (Vollrath et al., 1997; Liao et al., 2009) 等方式來調整築網的行為以減少風阻。這些可調整的築網行為具有一定程度之可塑性,即當風干擾存在或消失時,蜘蛛將採取不同的結網策略。除了降低風阻的行為具可塑性,由於風阻主要由大壺狀腺絲承受,推測大壺狀腺絲的性質對風干擾亦存在適應或可塑性。先前研究指出,在經過風干擾後,圓網蜘蛛所分泌的大壺狀腺絲具有較高之強度、韌度與延展性 (Liao et al., 2009; Wu et al., 2013),顯示蜘蛛可以感受風干擾程度並其大壺狀腺絲在生產的層次上也具有可塑性。
承上所述,蜘蛛絲面對風干擾之可塑性可以發生在蜘蛛絲分泌以至於結網策略不同等 2 個層面,特別是大壺狀腺絲的演化歷程在於克服捕捉飛行性昆蟲與避免風干擾的目的,兩者的演化方向似乎相似—加強絲的強度與韌度。大壺狀腺絲相對容易由蜘蛛活體直接收集,加上其扮演的重要功能且具有一定的可塑性,因此這類的蜘蛛絲仍是目前最被注目的蜘蛛絲研究,亦是仿生蜘蛛絲的主要模仿模型。然而,對於蜘蛛在分泌層次上如何調整蜘蛛絲性質尚屬推論而無直接證據,僅有間接證據顯示蜘蛛在有無接受風干擾後其大壺狀腺絲之胺基酸組成並未呈現明顯變化 (Liao et al., 2009)。
風中的工程師:二角塵蛛如何調整蜘蛛絲以抵抗風干擾
本研究以前人研究中對風干擾誘發大壺狀腺絲可塑性的二角塵蛛 (Cyclosa ginnaga) 為對象。在臺灣,二角塵蛛通常棲息於低海拔空曠無遮陰的微棲地,例如河堤或海岸。這些地區在其生長與繁殖季(3 至 9 月)皆有強烈的風干擾。將蜘蛛由野外連同其蜘蛛網一併取回實驗室後隨機分配至「風干擾組」與「無風組」,分別施予全天風干擾處理與無風干擾處理達 7 日。之後從網上取下蜘蛛再以相同速率拉出其大壺狀腺單絲,過程中蜘蛛被束縛在特定姿態且保持未受傷並清醒的狀態。每隻蜘蛛個體所抽取的絲樣本皆再隨機分成二組—「天然絲」與「超收縮絲」。天然絲會直接被微力拉伸儀測定其應力應變曲線,進而計算其強度、韌度、延展性及楊式係數等機械性質。而超收縮絲會先置入飽和蒸氣室中達到超收縮狀態後回復至乾燥再測量其機械性質。超收縮現象是由於蜘蛛絲內部的非結晶區域內之肽鏈間氫鍵被水分子打破後,使蜘蛛絲之微觀結構失去其特定排列方式,進而使蜘蛛絲「退化」至類似在腺體內初分泌的「未加工態」(Work, 1977; Liu et al., 2006; Dong et al., 2020) 。假若二角塵蛛可在分泌過程快速地調整蜘蛛絲的生產環境以達成絲機械性質之可塑性,我們可以預期,天然絲在受到不同風干擾的實驗處理之後,應如同前人研究般呈現差異,但超收縮絲則在不同風干擾處理之後,性質仍十分相似。
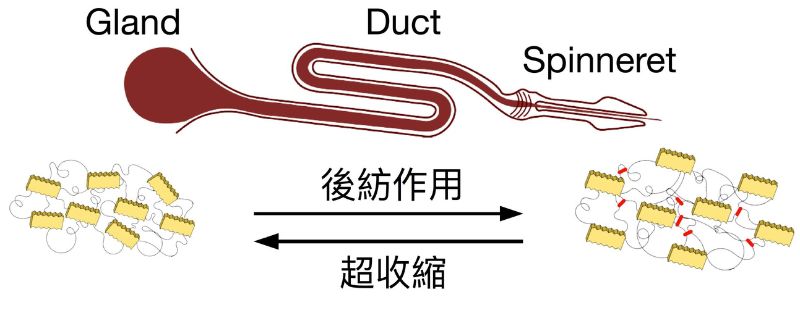
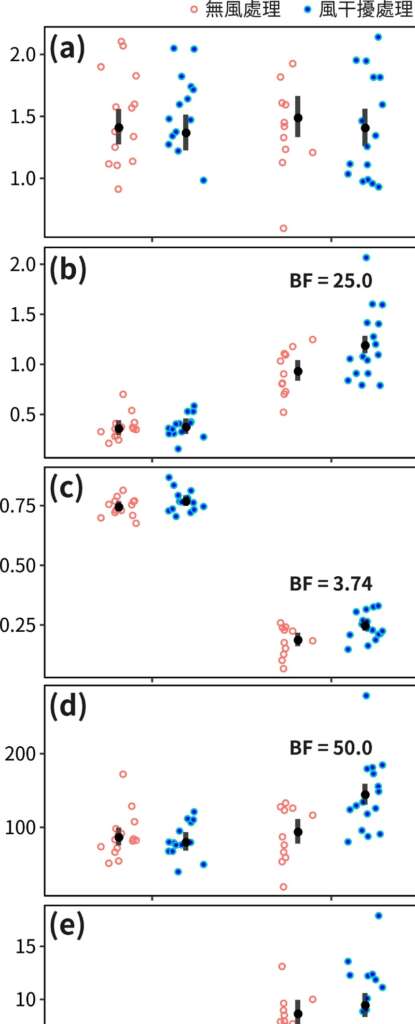
上述實驗之結果指出,二角塵蛛在經歷風干擾後,其大壺狀腺絲直徑並未改變,見下圖 a,但其強度、延展性及韌度則顯著提升 20-40% ,見下圖 b-d,而楊氏係數並未呈現明顯變化,見下圖 e。這與前人針對風干擾之研究成果十分相似。然而,比較超收縮絲的不同實驗處理,並無法偵測其間任何機械性質上的差異。此外,在比較超收縮絲的收縮量時亦發現風干擾組的收縮量明確但微幅大於無風處理組。以上結果證實,二角塵蛛在經歷風干擾後可藉由改變生產蜘蛛絲之體內微環境,但在不改變胺基酸組成的條件下,快速地調整大壺狀腺絲之機械性質,使絲產物及蜘蛛網更加堅韌因而達成降低風干擾的負面影響。這樣的可塑性主要作用是在非結晶區域產生更多氫鍵,使肽鏈摺疊地更加緊密。在大壺蜘蛛絲被拉伸的過程中,微觀結構中的非結晶結構首先受力,其中的肽鏈間氫鍵因絲延展而被迫斷裂,直到非結晶結構肽鏈延展至接近其極限。更多氫鍵曾使肽鏈摺疊更加緊密而有助於在拉伸過程中加強絲的延展性與韌度。本研究驗證了圓網蜘蛛藉由改變大壺狀腺絲的微觀排列方式,使蜘蛛絲機械性質有明確的提升,並指出未來針對蜘蛛絲腺體內微環境之監測將有助於吾人了解蜘蛛體內如何達成此種可塑性,並期待其成果可做為增強仿生蜘蛛絲性質的關鍵因子。


本著作由本館研究人員所提供,博學多文團隊編輯製作,以創用CC 姓名標示–非商業性– 禁止改作 4.0 國際 (CC BY-NC-ND 4.0) 授權條款釋出。若需要使用本篇的文字、圖像等,請洽本館出版室。


